

How Many Kinds of Dinosaurs Were There?
Tuntuu siltä, että vanhoina vanhoina hyvinä dinosaurusten renessanssia edeltävinä aikoina tieteen tuntemien dinosauruslajien määrä oli melko itsestään selvä asia. Oli Tyrannosaurus, Triceratops, Stegosaurus, Brontosaurus, ’Monoclonius’ ja kaikki muut klassikot. Se oli kuin fossiilisten eläinten baseball-joukkue. Tämä ei tietenkään koskaan pitänyt paikkaansa, sillä huolimatta siitä, että vain kymmenkunta eläintä näytti koskaan pääsevän eilisiin populaaritieteellisiin kirjoihin, oli satoja tuntemattomampia dinosauruksia, jotka eivät koskaan päässeet kovinkaan paljon parrasvaloihin ja jotka nimettiin samaan aikaan kuin tähtilajit.
Tänä päivänä, kun paleontologia on suositumpaa kuin koskaan ja kun ihmiset kehittyvät koko ajan paremmiksi ja paremmiksi paleontologian parissa, uusia dinosauruslajeja nimetään viikoittain kaikkialta maailmasta. Wikipedian mukaan muiden kuin aavisten dinosaurusten osalta meillä on tällä hetkellä noin 500 sukua, joka on eläimen tieteellisen nimen ensimmäinen ja kattavampi osa, kuten Canis koirille. Näiden sukujen sisällä on noin 1000 lajia. Se on tieteellisen nimen toinen ja eksklusiivisempi osa, kuten ”lupus” sanassa Canis lupus (susi) tai ”latrans” sanassa Canis latrans (kojootti). Tutkijat kiistelevät kuitenkin tästäkin luvusta, ja sitä, kuinka monta dinosauruslajia odottaa vielä maan alla löytymistään, on vaikea, ellei jopa mahdotonta sanoa. Näin ollen kysymykseen siitä, kuinka monta dinosauruslajia oli olemassa, on melko hankala vastata.
Uuden dinosaurussuvun tai -lajin löytämiseen on muutamia eri tapoja. Yksi tapa on mennä ja kaivaa esiin jotain ainutlaatuista, jota ei ole koskaan aiemmin löydetty. Voisi ajatella, että nykyään näin tapahtuu vain suhteellisen tutkimattomissa paikoissa maailmassa, joissa paleontologit vasta alkavat tutkia paikallisia kiviä, ja se on varmasti totta, että vähän tutkituilta alueilta löytyy uusia dinosauruslajeja. Albertan rajojen sisällä pitäytyen luoteisosan myöhäiskreettikautinen Wapiti-muodostuma, jossa Philip J. Currie -dinosaurusmuseo sijaitsee, voisi olla esimerkki tästä. Jopa kuuluisasti tunnetuilla alueilla, kuten provinssin eteläisellä puoliskolla, on kuitenkin edelleen aika ajoin aivan uusia dinosauruksia. Joskus löydämme uuden lajin jo olemassa olevasta suvusta, mutta joskus kyseessä on kokonaan uusi suku, joka antaa meille jännittävän uuden dinosauruksen nimen, josta voimme puhua. Uuden suvun tai lajin nimeäminen jostakin hiljattain löydetystä dinosauruksesta on aikaa vievä prosessi, sillä kun eläin on kerätty, se on valmisteltava, tutkittava huolellisesti, sille on tehtävä tiukka tieteellinen kuvaus, ja kun kuvaus on hyväksytty ja julkaistu, kaikki saavat nauttia uudesta dinosauruksesta. Huolimatta tekniikan ja teknologian kehittymisestä paleontologia etenee edelleen usein jäätävää vauhtia.
Meidän on myös muistettava se tosiasia, että maapallon tietyillä alueilla on geologisesti säilynyt vain tiettyjä aikaviipaleita. Meillä ei ole laajoja kalliopaljastumia jokaisesta iästä, ajanjaksosta ja aikakaudesta kaikista maailman paikoista, joista voisimme etsiä fossiileja. Tämä tarkoittaa, että geologisesta ajasta on kokonaisia lohkoja, joita emme ehkä koskaan pysty tutkimaan ja tietämään, mitä fossiilisia eläimiä kyseisellä ajanjaksolla eli. Kokonaiset dinosaurusten evoluution ja monimuotoisuuden vaiheet voivat olla ikuisesti arvoitus. Kaiken kukkuraksi emme voi olettaa, että kaikkien koskaan eläneiden dinosauruslajien jäsenet olisivat olleet niin onnekkaita, että niistä olisi saatu fossiileja. Fossiiliksi tuleminen edellyttää, että eläin on kuollut oikeaan aikaan ja oikeassa paikassa, jossa ympäristöolosuhteet ovat olleet sellaiset, että luut ja muut kovat osat ovat mineralisoituneet. Kova totuus on, että monet dinosaurukset elivät luultavasti ympäristöissä, jotka eivät olleet suotuisia fossiilisoitumisprosessille, ja siksi tiedot niiden koko olemassaolosta ovat kadonneet ikuisiksi ajoiksi. Eikö se olekin kauhean turhauttavaa? Voimme vain jatkaa kaivamista siellä, missä kiviä on, ja jatkaa toivomista.
Toinen tapa löytää uusi dinosauruslaji on etsiä museoiden tai yliopistojen kokoelmista jo kaivettuja esineitä. Paleontologian museoissa kaikkialla maailmassa on hyllyjä, laatikoita ja koteloita täynnä näytteitä, jotka odottavat analysointia ja kuvausta. Paleontologit ovat toki kunnianhimoisia ihmisiä, mutta he voivat tutkia vain tietyn määrän eläimiä kerralla. Usein tavarat, jotka ovat odottaneet perusteellista kuvausta, sysätään jatko-opiskelijoille opinnäytetöiksi. On väistämätöntä, että jotkut tunnistamattomat yksilöt osoittautuvat tutkittaessa jo todettujen lajien edustajiksi. Joskus osittainen luuranko tunnistetaan yhdeksi dinosaurustyypiksi ja myöhemmin tarkemman tarkastelun jälkeen uudelleen joksikin muuksi. Tästä julkaistiin hiljattain esimerkki, kun Kanadan luonnonmuseossa vuosikymmeniä hyvin yllättävästä kalliomuodostumasta kerätty nuori Daspletosaurus, jota luultiin vuosikymmeniä nuoreksi Daspletosaurukseksi, olikin mitä todennäköisimmin Albertosaurus, joka löytyi juuri silloin, kun sellaista odottaisi löytyvän (Mallon ym., 2019). Ja joskus fossiilit, joiden oletetaan olevan vakiintuneesta suvusta tai lajista, osoittautuvat joksikin aivan uudeksi. Yksi esimerkki tästä (vaikka niitä on paljon muitakin) on vuonna 2012 nimetty sarvipäinen dinosaurus Coronosaurus brinkmani, joka oli alun perin kuvattu seitsemän vuotta aiemmin Centrosaurus-suvun lajiksi (Ryan & Russel, 2005) (Ryan et al., 2012) (Ryan et al., 2012).
Kaikki tämä kertoo meille, että maassa tai museoissa piilossa on määrittelemätön määrä dinosaurusten sukuja ja lajeja odottamassa kuvausta ja nimeämistä. Uudet dinosaurukset eivät tule loppumaan lähiaikoina. On kuitenkin toinenkin syy olla epävarma dinosauruslajien määrästä, ja se on olemassa olevien sukujen ja lajien pätevyys, joka joidenkin mukaan ei ehkä olekaan niin varma. Taksonomiassa, eli eliöiden nimeämistä ja ryhmittelyä koskevassa tieteessä, on kaksi vastakkaista koulukuntaa. Toinen ryhmä ovat ”lumpers”, tutkijat, jotka painottavat enemmän hieman erilaisten eläinten yhtäläisyyksiä ja ryhmittelevät ne todennäköisemmin samaan sukuun tai lajiin. Toiseen ryhmään kuuluvat ”jakajat” – ne, jotka näkevät enemmän eroja eläinten välillä ja jakavat ne eri sukuihin tai lajeihin. ’Lumpkereita’ ja ’splittereitä’ esiintyy sekä nykyeläinten että sukupuuttoon kuolleiden eläinten taksonomiassa, ja tämä saa meidät miettimään kahdesti, miten nimeämme ja luokittelemme lajeja.
Lajit määritetään toisistaan eroaviksi tai samoiksi muutamien kriteerien perusteella. Näyttävätkö ne huomattavan erilaisilta, kuulostavatko ne erilaisilta tai käyttäytyvätkö ne eri tavalla? Onko niiden sisäinen anatomia ainutlaatuinen? Ehkä tärkein kysymys, joka on esitettävä, kun verrataan kahta samankaltaista mutta mahdollisesti erilaista eläintä, on se, voivatko ne tuottaa yhdessä täysin hedelmällisiä poikasia. Jos ne voivat, ne ovat todennäköisesti samaa lajia. Tämä on biologinen lajikäsitys, ja eläintieteilijät käyttävät sitä yleisenä nyrkkisääntönä määritellessään, ovatko eläimet samaa vai eri lajia (kasvitieteessä ja mikrobiologiassa se kuitenkin yleensä pettää). Eri lajien välinen risteytyminen on toki mahdollista, mutta se johtaa yleensä joko hedelmättömiin risteytyksiin (kuten hevosten ja aasien risteytyessä muuleiksi) tai jälkeläisiin, jotka eivät selviä hengissä syntymää pidemmälle, kuten vankeudessa elävien afrikkalaisten ja aasialaisten norsujen risteytysyrityksissä.

Pachyrhinosaurus (ylhäällä oikealla) ja Styracosaurus (alhaalla vasemmalla) näytteillä Philip J. Currie -dinosaurusmuseossa. Niiden kallot ovat melko erilaiset, mutta muut luut ovat melko samanlaiset. Mitä tästä pitäisi päätellä? Kuvat: Nicholas Carter
Sukujen tasolla asiat ovat myös monimutkaisia. Pitkään lajeja eroteltiin ulkonäön, käyttäytymisen ja sisäisen anatomian kaltaisten seikkojen avulla, mutta geneettisen sekvensoinnin suhteellisen tuoreen tehon ansiosta voimme ratkaista asioita entistä yksityiskohtaisemmin. Esimerkiksi noin puolet pohjoisamerikkalaisista ankoista luokiteltiin viime vuosikymmeninä Anas-sukuun. Viimeaikaiset molekyylianalyysit ovat kuitenkin osoittaneet, että tämä ei pidä paikkaansa (Gonzales ym., 2009), ja useat lajit jaettiin sukuihin, kuten Spatula, johon kuuluvat teerit ja lapasorsat, ja Mareca, johon kuuluvat sepelkyyhkyt ja lapasorsa (Gill & Donsker, 2017). Tämä ei tarkoita sitä, etteikö molekyylitodisteiden merkityksestä anatomisiin tietoihin nähden voisi kiistellä, mutta meillä on ainakin useita työkaluja käytettävissämme, joiden avulla voimme miettiä, miten luokitella nykyaikaisia eläimiä.
Kun on kyse fossiilisista eläimistä, käytettävissä on kuitenkin paljon vähemmän perusteita. Ilman molekyylitietoja sekä havaintoja risteytymisestä, käyttäytymisestä ja ulkonäöstä, ainoat todelliset työkalut, joita meillä on fossiilisten lajien luokitteluun, ovat anatomia ja suhteellinen ajoitus. Paleontologit analysoivat tarkasti sukupuuttoon kuolleiden eläinten, kuten dinosaurusten, luita ja keräävät tietoja niiden anatomisista piirteistä eli luonteenpiirteistä, joiden perusteella he voivat päätellä, onko kyseessä eri suku/laji vai ei. Kallioperä, josta fossiileja on löydetty, on myös informatiivinen, sillä on melko epätodennäköistä, että sama laji löytyy eri muodostumista, jotka edustavat eri aikavälejä, mutta saman suvun eri lajit voivat varmasti löytyä hieman erilaisista muodostumista. Esimerkiksi ankkaluokan dinosaurus Hypacrosaurus sternbergi löytyy Two Medicine -muodostumasta, kun taas Hypacrosaurus altispinus löytyy nuoremmasta Horseshoe Canyon -muodostumasta (Horner et al., 2004).
Vähän koko paleontologian historian ajan on käyty jatkuvaa keskustelua monien dinosaurus-sukujen ja -lajien pätevyydestä. Paleontologiset niputtajat näkevät enemmän yhtäläisyyksiä eri eläinten välillä ja puoltavat niiden ryhmittämistä harvempien nimien alle, kun taas jakajat väittävät päinvastoin. Dinosaurusten yhdistämistä tai jakamista koskeviin päätöksiin vaikuttaa paljon. Jotkin fossiilit saattavat olla epätäydellisiä ja vaikuttaa ainutlaatuisilta, vaikka ne eivät välttämättä ole sitä. Erillisten hampaiden perusteella nimettyjen epäilyttävien dinosaurus-sukujen moninaisuus osoittaa hyvin tämän päänvaivan. Vaihtoehtoisesti jotkin epätäydelliset yksilöt näyttävät ensi silmäyksellä kuuluvan jo vakiintuneisiin taksoneihin, mutta ne eivät ole riittävän täydellisiä, jotta asiasta voitaisiin olla varmoja. Royal Tyrrell Museumin mysteerinen Dinosaur Parkin pachyrhinosaurus on yksi tällainen esimerkki (Ryan ym., 2010).
Vuosien varrella tehdyt löydöt nuorista dinosauruksista sekoittavat myös taksonomian vesiä. Dinosaurusten kallot muuttuivat melko paljon eläinten kasvaessa, erityisesti mitä tulee niiden pään koristeisiin, kuten harjanteisiin, sarviin ja röyhelöihin. Olemme vasta oppimassa, miten radikaalisti dinosaurusten kallot muuttuivat kasvun aikana. Ainutlaatuisilta näyttäneiden lajien kallot tai kallonpalat on luokiteltu uudelleen muiden dinosaurusten nuoriksi tai ala-aikuisiksi eläimiksi. Nimet kuten ”Tetragonosaurus” (Evans ym., 2005), ”Monoclonius” (Ryan, 2006) ja ”Brachyceratops” (Ryan, 2007) on niputettu muihin sukuihin, joille ovat tyypillisiä aikuiset yksilöt.

Joitakin Pachyrhinosaurus lakustai -lajeissa nähtyjä hapsujen vaihteluita. Dinosaurukset eivät todellakaan olleet kaikki samanlaisia! Kirjoittanut Nicholas Carter
Olemme myös vasta alkaneet selvittää, kuinka paljon kehon muodon ja koon vaihtelu oli mahdollista saman lajin yksilöiden välillä dinosauruksilla. Paleontologit ja paleotaiteilijat ovat jo pitkään olleet tiukkoja rekonstruoimaan dinosauruksia melko kapeiden koko- ja muotolinjojen mukaan. Olemme kuitenkin selvittämässä, että aivan kuten saman lajin nykyaikaiset eläimet voivat saada erilaisia muotoja genetiikan, vammojen, mutaatioiden ja muiden vastaavien tekijöiden vuoksi, myös dinosaurukset saattoivat saada erilaisia muotoja. Philip J. Currie -dinosaurusmuseon ylpeys, ceratopsidi Pachyrhinosaurus lakustai, rekonstruoidaan taiteessa usein melko symmetrisellä hapsu- ja piikkijärjestyksellä – kaksi sisintä hapsun piikkiä eli epiossifikaatiota ovat pieniä ja sisäänpäin suuntautuvia, toinen pari on pitkä ja sivulle kaartuva ja loput ovat pieniä nystyröitä. Pipestone Creekin luukerrostumasta kaivettujen täytemorfologioiden villi kirjo osoittaa kuitenkin, että tämän dinosauruksen koristeita oli kaikenlaisia epäsymmetrisiä muotoja (Currie ym., 2008). Tuoreemmissa uutisissa uusi Styracosaurus-dinosauruksen täydellinen kallo on osoittanut, että myös tämän suvun kalloissa oli melko paljon vaihtelua ja epäsymmetriaa, ja kirjoittajat pitivät tätä hyvänä syynä niputtaa samankaltainen mutta arvoituksellinen Rubeosaurus Styracosauruksen kanssa samaan luokkaan (Holmes ym, 2019).
Joskus dinosaurustaksonien niputtamisen tai (useammin) jakamisen syynä on yksinkertaisesti tiettyjen paleontologien ego ja kilpailuasetelma. Tämä kävi selvimmin ilmi 1800-luvun lopun luusotien aikana, kun kilpailevat paleontologit O.C. Marsh ja E.D. Cope yrittivät tunnetusti haukkua toisiaan nimeämällä eniten lajeja ja mitätöimällä samalla toisen miehen lajit. Marsh esimerkiksi nimesi yli puoli tusinaa Triceratopsin lajia. Vain kahta niistä pidetään nykyään pätevinä. Cope nimesi itse kourallisen sukuja, jotka kaikki ovat melko hajanaisia ja jotka todennäköisesti kuuluvat Triceratopsille.
Jos toisaalta tarkastelee dinosaurussukuja, voi huomata, että useimmissa niistä on vain yksi tai kaksi lajia. Jos tätä verrataan elävien eläinten sukuihin, se on aika outoa. Pelkästään liskojen suku Varanus sisältää yli 50 elävää lajia, joiden koko ja muoto vaihtelee suuresti. Nykyaikaisista elossa olevista arkkosauruksista Crocodylus-suvussa on 14 lajia, ja nykyaikaisten lintujen suvuissa on yhdestä lajista useisiin kymmeniin lajeihin. Hyönteistiede puhaltaa tämän vesiperän, sillä joissakin hyönteissuvuissa on kussakin yli 100 lajia. Tämä on saanut jotkut kirjoittajat ehdottamaan, että dinosaurukset on jaettu liikaa ja että monet samankaltaiset suvut pitäisi oikeastaan niputtaa yhteen. Tämä on koskenut erityisesti sukuja, joilla on yleensä erinäköiset kallot mutta jotka ovat kaulasta alaspäin melko samanlaisia. Tämä paleontologien taipumus jakaa fossiilisten eläinten suvut kahtia johtuu usein varovaisesta ajattelutavasta – jos voi nimetä jotain, ei useinkaan kannata olettaa, että se on sama suku kuin jokin muu. Koon ja muodon vaihtelunsa vuoksi näyttää siltä, että dinosauruslajit arvostavat suuresti erottautumista toisistaan. Muilla biologian aloilla keskitytään kuitenkin yleensä yleisiin yhtäläisyyksiin. Hyönteistieteilijät todennäköisesti vieroksuisivat ajatusta siitä, että esimerkiksi ceratopsideihin kuuluu yli 40 erillistä sukua, joista useimpiin kuuluu vain yksi tai kaksi lajia.
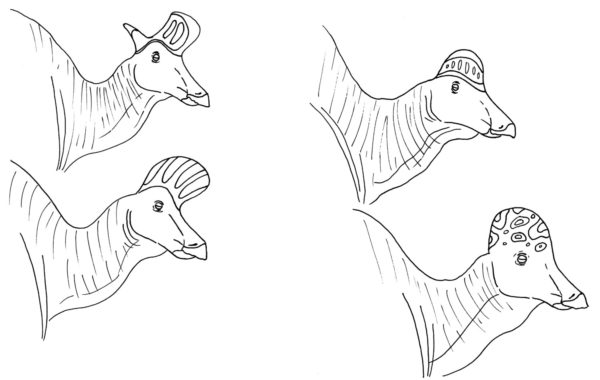
Kellon ympäri vasemmalta ylhäältä: Lambeosaurus lambei, Hypacrosaurus altispinus, Corythosaurus casuarius ja Lambeosaurus magnicristatus. Ovatko ne oikeasti kaikki samaa sukua? Onko sillä väliä? Kirjoittanut Nicholas Carter
Kirjailija ja taiteilija Gregory S. Paul on historiallisesti mennyt kiistanalaisiin linjoihin dinosaurusten yhdistämisessä. Väittäen, että monet toisiinsa liittyvät dinosaurus-suvut olivat kallojensa ulkopuolella erottamattomia, Paul on vuosien varrella yrittänyt synonyymisoida monia dinosaurus-sukuja. Tunnettuja tapauksia ovat esimerkiksi Dale Russellin ehdotus, jonka mukaan tyrannosaurukset Albertosaurus ja Gorgosaurus ovat samaa sukua (Russell, 1970), mistä useimmat nykyaikaiset tyrannosaurusasiantuntijat eivät ole samaa mieltä, sekä Velociraptorin ja Deinonychuksen synonyyminen, joka on nyt hylätty (Paul, 1988). Paul on myös väittänyt esimerkiksi, että lambeosauriinit Corythosaurus, Lambeosaurus ja Velafrons pitäisi yhdistää Hypacrosaurukseen ja että suuri osa centrosaurineista pitäisi yhdistää Centrosaurukseen (Paul, 2016). Tohtori Darren Naish kertoo tästä lisää täällä. Jos nämä esimerkit (ja monet muut Paulin väittämät) olisivat laajalti paleontologien hyväksymiä, kelvollisten dinosaurus-sukujen määrä olisi huomattavan pieni. Näin laaja taksonominen uudistaminen vaatisi kuitenkin valtavan määrän tutkimusta ja tietoja, jotta se saisi tieteellistä tukea. Subjektiiviset havainnot eri eläinten samankaltaisuudesta kaulan alapuolella eivät ole kovaa näyttöä. On kuitenkin mielenkiintoista huomata, että Paul on myös ehdottanut joitakin taksonomisia jakoja dinosaurussukuihin, kuten Giraffatitan Brachiosauruksesta (Paul, 1988) ja Mantellisaurus Iguanodonista (Paul, 2008), jotka ovat sittemmin saaneet tukea tiedemiehiltä (Taylor, 2009) (Norman, 2010).
Jos nyt alat jo miettiä, mikä ylipäätään enää muodostaa dinosaurussuvun, et ole yksin. Lisäksi, jos mietit, onko sillä ylipäätään edes niin paljon väliä, et ole myöskään yksin. Dinosaurusten nimeäminen ja luokittelu on hyödyllinen tapa tutkia ja ymmärtää niitä tieteellisesti. Me ihmiset rakastamme lajitella, luokitella ja laskea asioita. Onko sillä kuitenkaan mitään väliä, jos esimerkiksi Eotriceratops xerinsularis osoittautuisi vain Triceratops-suvun varhaiseksi lajiksi? Muuttuuko mikään todella? Ei oikeastaan. Paleontologit ja muut dinosaurusnörtit (kuten minä) rakastavat väitellä nimien ja luokitusten kaltaisista yksityiskohdista, mutta ne kaikki ovat ihmisen luomuksia kuvaamaan eläviä olentoja tietyssä pisteessä evoluution spektrissä.
Dinosaurustyyppien määrä ei siis ole varma, nouseva luku. Pikemminkin se riippuu osittain siitä, kenen kanssa puhut ja mitä todisteita arvostat. Kaikesta väittelystä huolimatta paleontologit kuitenkin löytävät edelleen uusia lajeja, ja se on aina jännittävää.
Toimittaja Nicholas Carter

Currie, P.J., Langston, W., ja Tanke, D.H. (2008). ”A new species of Pachyrhinosaurus (Dinosauria, Ceratopsidae) from the Upper Cretaceous of Alberta, Canada.” pp. 1-108. In:
Currie, P.J., Langston, W., and Tanke, D.H. 2008. A New Horned Dinosaur from an Upper Cretaceous Bone Bed in Alberta. NRC Research Press, Ottawa, Ontario, Kanada. 144 pp.
Evans, David C.; Forster, Catherine F.; Reisz, Robert R. (2005). ”The type specimen of Tetragonosaurus erectofrons (Ornithischia: Hadrosauridae) and the identification of juvenile lambeosaurines”. In Currie, Phillip J.; Koppelhus, Eva (eds.). Dinosaur Provincial Park: A Spectacular Ancient Ecosystem Revealed. Bloomington: Indiana University Press. pp. 349-366.
Gill, Frank; Donsker, David, eds. (2017). ”Huutelijat, ankat, hanhet & joutsenet”. Maailman lintuluettelon versio 7.3. International Ornithologists’ Union. Haettu 23.7.2017.
Gonzalez, J.; Düttmann, H.; Wink, M. (2009). ”Phylogenetic relationships based on two mitochondrial genes and hybridization patterns in Anatidae”. Journal of Zoology. 279: 310-318.
Robert.B. Holmes; Walter Scott Persons; Baltej Singh Rupal; Ahmed Jawad Qureshi; Philip J. Currie (2019). ”Morfologinen vaihtelu ja epäsymmetrinen kehitys Styracosaurus albertensiksen kallossa”. Cretaceous Research. in press: Article 104308. doi:10.1016/j.cretres.2019.104308.
Horner, John R.; Weishampel, David B.; Forster, Catherine A (2004). ”Hadrosauridae”. In Weishampel, David B.; Dodson, Peter; Osmólska Halszka (eds.). The Dinosauria (2. painos). Berkeley: University of California Press. pp. 438-463.
Mallon, J.C., Bura, J.R., Schumann, D. and Currie, P.J. (2019), A Problematic Tyrannosaurid (Dinosauria: Theropoda) Skeleton and Its Implications for Tyrannosaurid Diversity in the Horseshoe Canyon Formation (Upper Cretaceous) of Alberta. Anat Rec. doi:10.1002/ar.24199
Norman, David. (2010). ”A taxonomy of iguanodontian (Dinosauria: Ornithopoda) from the lower Wealden Group (Cretaceous: Valanginian) of the southern England” (PDF). Zootaxa. 2489: 47-66.
Paul, Gregory S. (1988). Predatory Dinosaurs of the World. New York: Simon & Schuster. s. 464. ISBN 978-0-671-61946-6.
Paul, G.S. (1988). ”The brachiosaur giants of the Morrison and Tendaguru with a description of a new subgenus, Giraffatitan, and a comparison of the world’s largest dinosaurs”. Hunteria. 2 (3): 1-14.
Paul, Gregory S. (2008). ”A revised t
axonomy of the iguanodont dinosaur generaures and species”. Cretaceous Research. 29 (2): 192-216.
Paul, Gregory S. (2016). The Princeton Field Guide to Dinosaurs 2nd Edition. United States of America: Princeton University Press.
Russell, Dale A. (1970). ”Tyrannosaurukset Länsi-Kanadan myöhäisestä liitukaudesta”. National Museum of Natural Sciences Publications in Paleontology. 1: 1-34.
Ryan, M.J. (2006). ”The status of the problematic taxon Monoclonius (Ornithischia: Ceratopsidae) and the recognition of adult-sized dinosaur taxa”. Geological Society of America Abstracts with Programs. 38 (4): 62.
Ryan, M. J.; Russell, A. P. (2005). ”A new centrosaurine ceratopsid from the Oldman Formation of Alberta and its implications for centrosaurine taxonomy and systemics”. Canadian Journal of Earth Sciences. 42 (7): 1369. doi:10.1139/e05-029
Ryan, Michael J.; Holmes, Robert; Russell, A.P. (2007). ”A revision of the late Campanian centrosaurine ceratopsid genus Styracosaurus from the Western Interior of North America” (PDF). Journal of Vertebrate Paleontology. 27 (4): 944-962.
Ryan, M. J.; Eberth, D. A.; Brinkman, D. B.; Currie, P. J.; Tanke, D. H., ”A New Pachyrhinosaurus-Like Ceratopsid from the Upper Dinosaur Park Formation (Late Campanian) of Southern Alberta, Canada”, teoksessa Ryan, M. J.; Chinnery-Allgeier, B. J.; Eberth, D. A., toim. (2010), New Perspectives on Horned Dinosaurs: The Royal Tyrrell Museum Ceratopsian Symposium, Indiana University Press
Ryan, M. J.; Evans, D. C.; Shepherd, K. M.; Sues, H. (2012). ”A new ceratopsid from the Foremost Formation (middle Campanian) of Alberta”. Canadian Journal of Earth Sciences. 49 (10): 1251. doi:10.1139/e2012-056
Taylor, M.P. (2009). ”A Re-evaluation of Brachiosaurus altithorax Riggs 1903 (Dinosauria, Sauropod) and its generic separation from Giraffatitan brancai (Janensch 1914)” (PDF). Journal of Vertebrate Paleontology. 29 (3): 787-806.