
Introduktion av cyklin
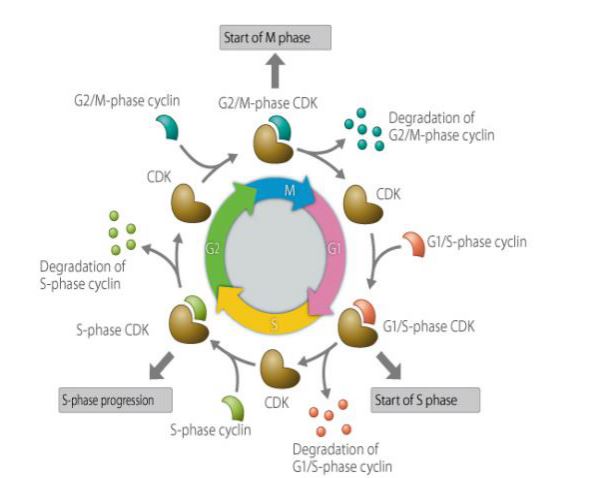
Cyklin är en typ av protein som uttrycks, ackumuleras och bryts ner i en cellcykel, och det interagerar med cyklinberoende kinaser för att påverka cellcykelfunktionen. Cyklin har medverkat i cellcykelreglerade proteiner, och dess koncentration är cyklisk och cyklisk i cellcykeln. Beroende på cellcykelns stadium är koncentrationen ibland så hög som flera tusen gånger och ibland sjunker den till noll. Som en reglerande underenhet binder cyklin till och aktiverar cyklinberoende proteinkinaser. Olika cykliner uttrycks under olika perioder i cellcykeln. I däggdjursceller börjar cyklin A uttrycka sig och ackumuleras gradvis i den tidiga G1-fasen och når G1/S-övergången, och innehållet når sitt maximum och kvarstår i G2/M-fasen. Cyklin B uttrycks från den sena G1-fasen och ackumuleras gradvis. Det når sitt maximum i den sena G2-fasen och bibehålls till mitten av M-fasen, för att sedan snabbt brytas ned. Cyklin D, som är ett cyklin i G1-fasen, uttrycks kontinuerligt i cellcykeln, medan cyklin E börjar uttrycka sig och gradvis ackumuleras i den sena M-fasen och den tidiga G1-fasen, når sitt maximum i den sena G1-fasen, för att sedan gradvis nå sitt maximala innehåll och avta fram till den sena G2-fasen. Cyklin spelar en viktig roll i regleringen av cellcykeln. Att studera cyklins mekanism har stor betydelse för behandlingen av cancer.
Cyklinfamiljemedlemmar och deras funktioner respektive
Hanskt cyklin har hittills isolerats och namngivits i sammanlagt 8 kategorier, nämligen A till H, och innefattar vissa underklasser (t.ex. cyklin D1, D2, D3), av vilka F, G, H är de senaste upptäckarna. Alla cykliner har vissa likheter i aminosyrasekvensen och fungerar därför som molekylära strukturmarkörer för cykliner. Denna homologa aminosyraregion i cykliner är den så kallade cyklinkassetten, som är en mycket bevarad sekvensregion på 100-150 aminosyror mellan cykliner från olika organismer. Det mänskliga cyklinet innehåller också en defekt kassett eller en aminosyrasekvensregion som är rik på prolin-, glutaminsyra-, asparaginsyra-, serin- och treoninrester (nedan kallad PEST-aminosyrasekvensregionen). Cyklin B:s bindning till det cyklinberoende kinaset p34CDC2 kontrollerar in och utgången av mitos, som syntetiseras från S-fasen till G2/M-fasen och gradvis ackumuleras och binds till p34CDC2. I M-fasen bryts den successivt ned; att lämna M-fasen beror tvärtom på den snabba nedbrytningen av cyklinet i det sena delningsstadiet, vilket resulterar i att den inaktiva underenheten p34CDC2 frigörs. I mänskliga celler ackumuleras cyklin B i cytoplasman och går in i kärnan innan kärnmembranet försvinner. När cyklin B/CDC väl är aktiverat fosforylerar det en grupp proteiner, bland annat lamin, vimentin och kalcium, vilket spelar en viktig roll för kromosomkoncentrationen, kärnmembranets upplösning, depolymerisering av intermediära filament och reorganisering av mikrofilament. Cyklin B spelar också en viktig roll i mitosens fasövergång, vilket är ett klassiskt klyvningscykelprotein. Cyklin A spelar en roll i både G2/M- och G1/S-fasövergångarna. Det uppträder före starten av DNA-syntesen, och det ökar gradvis fram till förklyvningsstadiet och bryts ned i mellanstadiet. När det inducerar mitos skiljer det sig från det äkta mitotiska cyklin B i många avseenden. Cyklin C-innehållet förändras föga under hela cellcykeln, med endast en liten ökning i tidigt G1. Cyklin C mRNA toppar i mitten av G1 i synkroniserade HeLa-celler, vilket är tidigare än cyklin A, som kan ha en roll i G1. Cyklin E har ett distinkt periodiskt uttryck i cellcykeln, och dess topp ligger i övergången mellan G1 och S-fasen. Den har funktionen att kontrollera att cellen går in i S-fasen, vilket kan vara den hastighetsbegränsande faktorn i G1/S-fasövergången. I däggdjursceller binder det liksom cyklin A till p33CDK2, ett komplex som har histon H1-kinasaktivitet i tidig G1- och S-fas, men cyklin A och E kan interagera med retinoblastomprotein för att separera komplex, vilket tyder på att CDK2/cyklin A eller E-komplex indirekt kan reglera genuttrycket i G1- och S-faser. Cyklin E kan spela en roll för cyklin D, som spelar en viktig roll för initiering av DNA-replikation, G1/S-konvertering, och dess överuttryck påskyndar cellens inträde i S-fasen. Cyklin D kan kontrollera den tidiga G1-fasen i processen, antingen före eller samtidigt med cyklin E. Cyklin D har tre subtyper (D1, D2, D3), och dess uttryck varierar från cell till cell och styrs av olika kromosomala regioner (D1: 11q13; D2: 12p13; D3: 6p21). Subtyperna kan ha olika effekter. Cyklin D:s anmärkningsvärda egenskaper är att det induceras att uttrycka sig som svar på externa stimuli, visar hög inducerbarhet för tillväxtfaktorer och fungerar som en tillväxtfaktorsensor, som spelar en viktig roll när det gäller att koppla samman externa signaler med den inre cellcykeln. Rollen, i sin tur, härleder dess okontrollerade uttryck gör att cellcykeln inte längre eller mindre beroende av tillväxtfaktorer och kan framkalla cancer. Cyklin D-CDK-komplexet är den bästa kandidaten för G1-stadiet retinoblastoma (Rb)-proteinkinas. Cyklin D binder till den N-terminala regionen av Rb-proteinet, fosforylerar Rb i den sena G1-fasen, och cyklin D1-syntes och aktivering leder till inaktivering av Rb-fosforylering och nedreglering av cyklin D1, och D1-CDK4-komplexet bildar en negativ återkoppling i G1-fasen, vilket i sin tur stänger av uttrycket av cyklin D1. Cyklin D och Rb spelar en viktig roll för cellproliferation och differentiering, när cyklin D1 är annorlunda i G1-fasen. I källuttrycket är Rb-fosforyleringen tidigare än normalt; G1-fasen påskyndas, och anticyklin D1-antikropp mikroinjiceras i G1 tidigt till metafasceller; de flesta cellerna stoppas före S-fasen, och avsaknad av funktionellt Rb kan inte orsaka detta block; man drar slutsatsen att cyklin D1:s viktiga roll är att inaktivera fosforyleringen av Rb, och därigenom inducera inträde i S-fasen och DNA-replikation. CyklinF är det vanligaste cyklinproteinet (molekylvikt 87 kDa), och dess mRNA uttrycks universellt i olika mänskliga vävnadsceller, och det finns betydande förändringar i cellcykeln. Toppen ligger i G2-fasen, liksom cyklin A, och minskar innan cyklin B:s mRNA-nivå sjunker. Cyklin F-proteinet ackumuleras i den intercellulära fasen och förstörs under mitosfasen. Det finns i kärnan i de flesta celler. Övexpression eller mutation i mänskliga celler resulterar i en brist på PEST-aminosyrasekvensregioner vilket leder till en ökning av celler i G2-fasen. Cyklin G liknar mest Cyklin B från fissionsjäst, som spelar en roll i G1/S-omvandlingen, men mRNA av cyklin G har inget tydligt cellberoende men kan stimuleras av celltillväxt. Faktorn inducerar och upprätthåller en förhöjd nivå. Cyklin g-genen innehåller två olika p53-bindningsställen, varav det ena ligger uppströms transkriptionsinitieringsstället, vilket tyder på att p53 har potential att effektivt aktivera cyklin g-genen. Cyklin H, som är en nedströmsmediator till p53 åtminstone i biologisk effekt, är ett protein med en molekylvikt på 37 kD som hittats genom isolering och rening av CDK-aktiverande kinas (CAK).
Funktionen hos cyklin
Studien av förhållandet mellan cellcykel och cancer har vägledande betydelse för klinisk onkologisk forskning. Den kan ge vissa mål för klinisk behandling, t.ex. blockering av cyklin D, eller efterlikna verkan av cellcykelhämmare för att hämma cancercelldelningen, och ge vissa indikatorer för klinisk diagnos, differentialdiagnos och prognos. Michaelet al analyserade bcl-1 genrearrangemanget och cyklin D1-proteinuttrycket i 32 fall av mantelcellslymfom, 17 fall av bcl-1 genrearrangemang och 24 fall av cytoplasmatiskt cyklin D1-uttryck, och 40 fall av kontrollgrupp endast 9 fall av icke-MCL B-cellslymfom var positiva. De anser att cyklin D1 kan användas som en differentialdiagnos av MCL och en differentialdiagnos av B-cellslymfom. Cyklin uttrycks periodiskt i cellcykeln och vissa specifika cyklin-CDK-komplex krävs för passage genom en viss cellcykel, vilket tyder på att cyklin kan användas som en indikator på cellernas proliferativa tillstånd. Uttryckskvoten för ett visst cyklin kan förutsäga andelen celler i en viss vävnad som befinner sig i en viss cellcykel. Ju högre malignitet hos tumörcellerna, desto allvarligare är cellcykelstörningen, så det är möjligt att spegla patienternas prognos genom den information som visas i cellcykeln, och fungera som en möjlig prognostisk indikator. Keyomarsi et al. använde 9 fall av kirurgiska prover från bröstcancer och intilliggande icke-cancerösa vävnader som kontrollstudier: I 8 fall av cancervävnad var cyklin E onormalt uttryckt i kvalitet och kvantitet, och det fanns minst 3 olika molekylvikter. Cyklin E var överuttryckt, medan c-erbB2 var överuttryckt i endast 3 fall. Med ökningen av tumörstadiet ökade innehållet av cyklin E-protein betydligt, medan nivån av proliferating cell nuclear antigen (PCNA) endast ökade svagt. Cyklin E-strukturen observerades i 4 av de högsta stadieindelningarna. Dutta et al. använde antikroppar mot cyklin A, B och E för att upptäcka 48 paraffinsklipp av bröstcancer och fann att det genomsnittliga positiva indexet för cyklin A och B ökade signifikant med ökningen av S-fasen (P < 0,05). Cyklin A var positivt korrelerat med Ki-67 och med S-fasfraktionen (P < 0,05). Bellacosaet al studerade 51 fall av primärt laryngealt skivepitelcancer och följde upp i 29 månader. Man tror att cyklin-genamplifiering kan användas som en oberoende prognostisk indikator för larynxcancer. Bettiche et al. studerade dock 53 fall av icke-småcellig lungcancer med kirurgisk resektion, och 25 fall hade överexpression av cyklin D1, vilket var förknippat med dålig vävnadsdifferentiering, mindre lymfocytär infiltration i tumören och lägre lokal recidivfrekvens (P < 0. 05). De anser att det behövs större kliniska studier för att ytterligare testa den prognostiska betydelsen av cyklin D1-uttryck.
Referens
- He G, Kuang J, Koomen J, et al. Rekrytering av trimeriskt proliferating cell nuclear antigen av G1-fas cyklinberoende kinaser efter DNA-skada med platinabaserade antitumörmedel. British Journal of Cancer. 2013, 109(9):2378-2388.
- Gang W, Cao J, Chen P, et al. Temporal and Spatial Expression of Cyclin H in Rat Spinal Cord Injury. Neuromolekylär medicin. 2011, 13(3):187-196.
- Ibrahim N, Nazimi A J, Ajura A J, et al. De kliniska egenskaperna och uttrycket av bcl-2, Cyclin D1, p53 och Proliferating Cell Nuclear Antigen i syndromiska och icke-syndromiska keratocystiska odontogena tumörer. Journal of Craniofacial Surgery. 2016, 27(5):1.
- Wang X, Wolgemuth D J, Baxi L V. Overexpression of HOXB5, cyclin D1 and PCNA in congenital cystic adenomatoid malformation. Fosterdiagnostik & Terapi. 2011, 29(4):315-320.