
Um modelo é definido no Novo Dicionário Colegial de 1978 do Webster como uma molécula (como o RNA) num sistema biológico que carrega o código genético de outra molécula. Além disso, o Dicionário Oxford Conciso de 1995, Nona Edição, descreve um modelo como o padrão molecular que rege a montagem de uma proteína, etc. De acordo com estas definições amplas, DNA é o modelo para DNA (em replicação de DNA) e para RNA (em transcrição ), e RNA é o modelo para proteína (em tradução ). Em outras palavras, um modelo é uma entidade molecular essencial para a transferência de informação genética do DNA para o DNA e do RNA para a proteína (o dogma central). Mais precisamente, a palavra “modelo” é usada quando uma molécula transferindo informação (o modelo) está em contato direto com a molécula receptora (o produto). Portanto, neste sentido, o RNA não é um modelo para proteína porque não ocorre interação direta entre o RNA mensageiro e a proteína durante a tradução (ver biossíntese de proteína). Nos casos que são a exceção ao dogma central, o RNA viral é sintetizado a partir do RNA usando o RNA como modelo (ver vírus RNA) e, além disso, o DNA é sintetizado usando o RNA como modelo durante a infecção por retrovírus (ver também Transcrição reversa).
Na replicação de DNA, a hélice dupla é desenrolada, e cada molécula de DNA de cadeia única é usada como um modelo para sintetizar uma cadeia complementar. Como o DNA sofre replicação semi-conservadora, o DNA parental serve como um modelo e torna-se um componente da molécula filha de DNA. Na transcrição, em contraste, uma fita de DNA serve como um modelo para a síntese de RNA que depois se dissocia do modelo (ver Transcrição). As polimerases de DNA que montam desoxirribonucleotídeos na cadeia de DNA do modelo de acordo com a Adenina (A):Tiamina (T), Guanina(G):Citosina (C) regra complementar base (emparelhamento base Watson-Crick) requerem tanto um modelo como um primer para que a reação ocorra (Fig. 1). Consequentemente, a síntese de DNA é precedida pela síntese de pequenas moléculas de RNA com menos de 10 bp por RNA primase, que pode iniciar a síntese de RNA complementar ao DNA modelo (ver fragmentos de Okazaki). O RNA primer é eventualmente substituído pelo DNA através do cordão de DNA retardado estendido da região que precede a síntese do RNA primer (veja Garfo de Replicação (Intermediário Y-Fork)). A relação modelo-produto através da complementaridade base de A a T e G a C não é 100% precisa, mas resulta na formação de pares errados uma vez a cada 10 3 a 104 vezes. Por outro lado, os desajustes geralmente ocorrem apenas uma vez em 10 a 10 quando os genomas são replicados dentro da célula . Isto é conseguido pela actividade de revisão das próprias polimerases do ADN e por outros mecanismos de reparação do ADN.
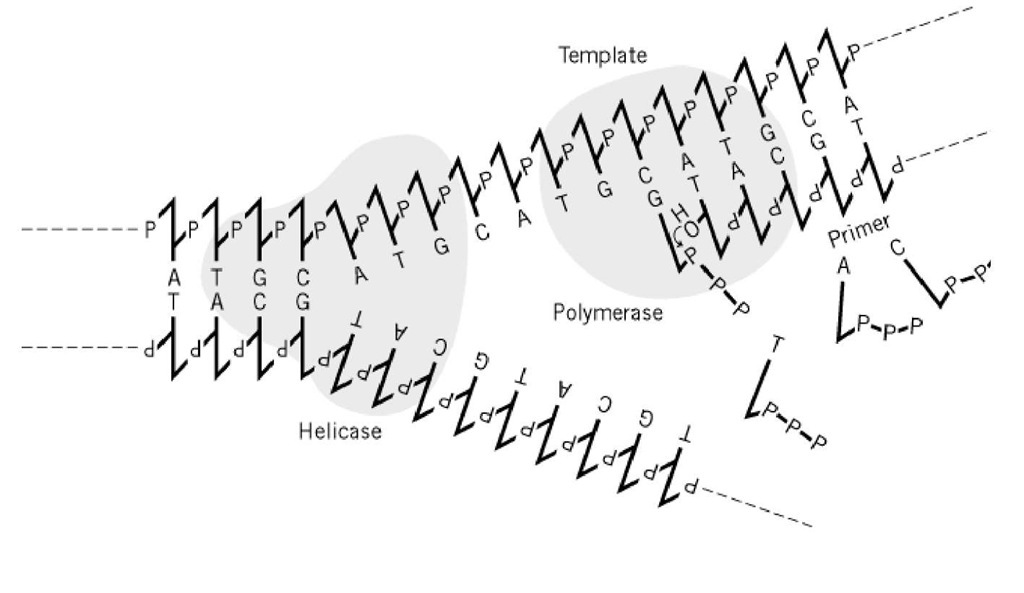
Figure 1. A DNA polimerase requer modelo e primer. Em um garfo de replicação crescente, fios únicos de DNA fornecidos por DNA helicase servem como templos para DNA polimerases. As enzimas sintetizam ligações fosfodiésteres entre o 3′( fim do primer, fio recentemente sintetizado, e o desoxirribonucleotídeo-trifosfato que é de base de reparo correto com fio de modelo. Apenas a síntese da cadeia principal é mostrada esquematicamente aqui.
A relação modelo-produto é muito importante na transferência de informação genética em sistemas biológicos. Contudo, a natureza físico-química da relação em si não é suficientemente precisa para realizar os processos biológicos regulados com precisão, e muitas proteínas foram desenvolvidas para reparar os erros que inevitavelmente resultam.