
A lizoszómák gömb alakú, membránhoz kötött organellumok, amelyeket a golgi-készülék hoz létre. Hidrolitikus enzimeket tartalmaznak, és így a sejt újrahasznosító rendszerének részeként működnek.
Ebben a cikkben a lizoszómák szerkezetét, szintézisét és működését vizsgáljuk meg, és megvizsgáljuk jelentőségüket a klinikai gyakorlatban.
Szerkezet
A lizoszómák a sejtekben található savas membránhoz kötött, általában körülbelül 1 mikrométer hosszúságú organellumok. A lizoszómák számos hidrolitikus enzimet tartalmaznak, amelyek hidrolízisreakciókat katalizálnak.
A lizoszómát körülvevő membrán létfontosságú, hogy ezek az enzimek ne szivárogjanak ki a citoplazmába és ne károsítsák a sejtet belülről. A lizoszóma savas pH-értékének fenntartása érdekében a lizoszómamembránon keresztül protonok aktívan szállítódnak az organellumba.
Szintézis
A lizoszóma és a benne lévő enzimek külön-külön szintetizálódnak. A lizoszomális fehérjék ugyanúgy képződnek, mint bármely más fehérje. Az első lépés az mRNS-szál előállításának beindítása a megfelelő DNS-szakaszokból. Az mRNS-szálak továbbhaladnak a durva endoplazmatikus retikulumba, ahol a riboszómák felépítik a hidrolitikus enzimeket.
Fontos, hogy ezeket a Golgi-apparátusban mannóz-6-foszfáttal jelölik meg, hogy a lizoszómába irányítsák őket. Ennek eredményeképpen az ezeket az enzimeket tartalmazó vezikulák a Golgi-apparátusból bomlanak ki. Két enzim felelős a mannóz-6-foszfát tag rögzítéséért: Az N-acetilglükozamin-foszfotranszferáz és az N-acetilglükozamin-foszfoglükozidáz.
Ez az immár a citoplazmában lévő vezikulum ezután egy késői endoszómához kötődik, amely egy másik savas, membránhoz kötött organellum. A késői endoszóma membránjában protonpumpa található, amely savasan tartja belső környezetét. Az alacsony pH a fehérje disszociációját okozza a mannóz-6-foszfát receptorról. Ez a receptor ezután visszakerülhet a Golgi-apparátusba.
A foszfátcsoportot a mannóz-6-foszfát-tagról is eltávolítják, hogy megakadályozzák a teljes fehérje visszatérését a Golgi-apparátusba. A késői endoszóma végül lizoszómává érhet, miután megkapta az enzimeket a Golgi-apparátusból.
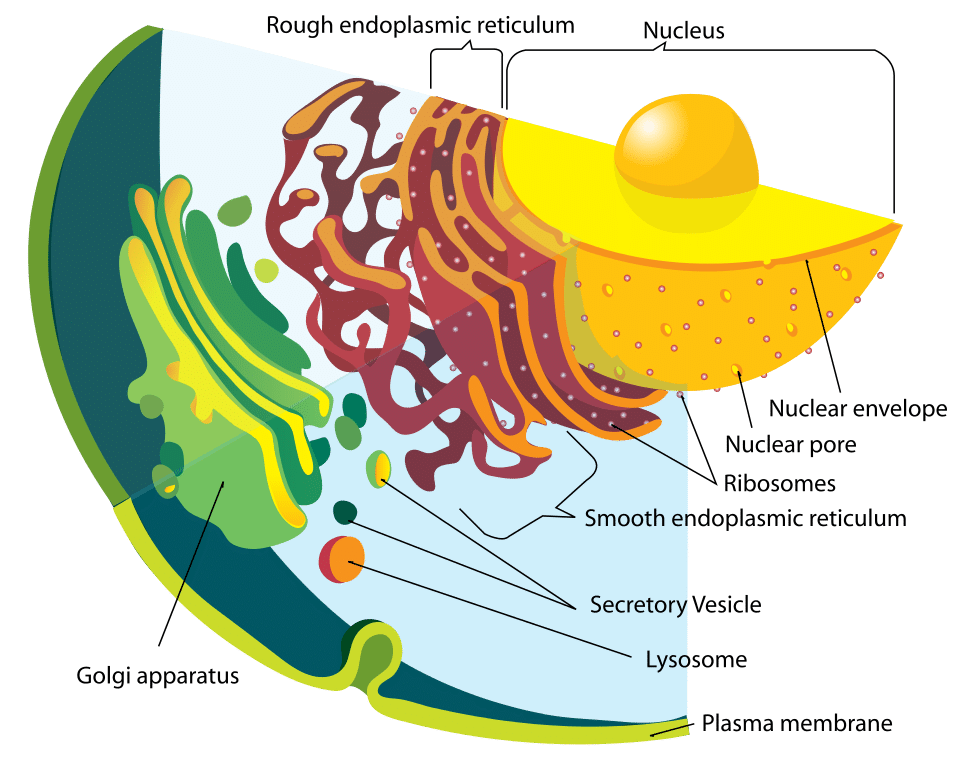 1. ábra – Az endomembrán rendszer ábrája
1. ábra – Az endomembrán rendszer ábrájaFunkció
A lizoszómában található hidrolitikus enzimek lehetővé teszik az idegen részecskék megsemmisítését. A lizoszómák fontos szerepet játszanak a fagocitózisban. Amikor a makrofágok idegen részecskéket fagocitálnak, azokat egy fagoszómába zárják. A fagoszóma ezután egy lizoszómához kötődik, hogy fagoliszoszómát alkosson.
Ezek az enzimek kritikus szerepet játszanak az oxigénfüggetlen pusztító mechanizmusokban. A lizoszómák az endocitózison keresztül történő kórokozó bejutás elleni védekezést is segítik azáltal, hogy lebontják a kórokozókat, mielőtt azok elérnék a citoplazmát.
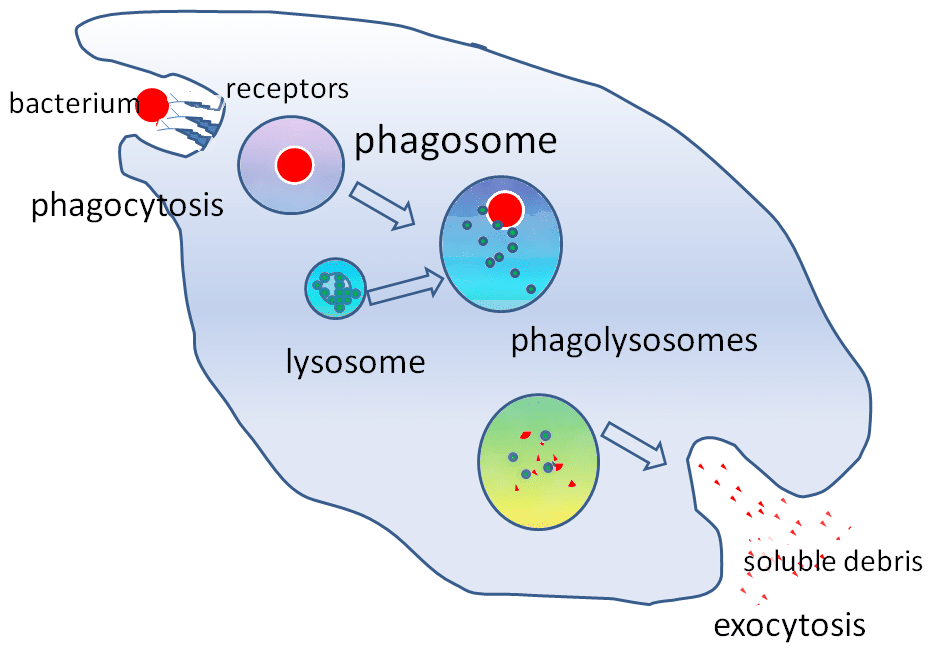 2. ábra – A lizoszóma szerepe a fagocitózisban
2. ábra – A lizoszóma szerepe a fagocitózisbanKlinikai jelentőség
I-sejtes betegség
Ezt az N-acetilglükozamin-foszfotranszferáz enzim genetikai hibája okozza. Ez az enzim létfontosságú a mannóz-6-foszfát hozzáadásához a lizoszóma célú fehérjékhez. Ez azt eredményezi, hogy a lizoszómális enzimek nem megfelelően célzottan működnek. Ennek eredményeként mind a vizeletben, mind a véráramban jelentős mennyiség található.
Lizoszomális tárolási betegségek
Ezek a lizoszómákat érintő genetikai állapotok egy csoportja. Az állapotok a tünetek, a tünetek és a betegek demográfiai jellemzői tekintetében nagymértékben eltérnek egymástól. Többféle osztályozás létezik; a leggyakoribb ilyen állapot a Gaucher-kór. Ezt a béta-glükocerebrosidáz hiánya okozza. Ez az enzim a glükocerebrozid lebontásához szükséges. Ezen enzim nélkül a glükocerebrozid képes felhalmozódni a sejtekben, ami károsíthatja azokat. A tünetek közé tartozik a hepatosplenomegália és a vérszegénység.