
Malli määritellään vuoden 1978 Webster’s New Collegiate Dictionary -sanakirjassa molekyyliksi (kuten RNA) biologisessa järjestelmässä, joka kantaa geneettistä koodia toiselle molekyylille. Myös vuoden 1995 Concise Oxford Dictionary, Ninth Edition kuvaa templaattia molekyylimallina, joka ohjaa proteiinin kokoamista jne. Näiden laajojen määritelmien mukaan DNA on malli DNA:lle (DNA:n replikaatiossa) ja RNA:lle (transkriptiossa ), ja RNA on malli proteiinille (translaatiossa ). Toisin sanoen templaatti on molekyylikokonaisuus, joka on välttämätön geneettisen informaation siirtymiselle DNA:lta DNA:lle RNA:lle proteiiniksi (keskeinen dogmi). Tarkemmin sanottuna sanaa ”templaatti” käytetään silloin, kun informaatiota siirtävä molekyyli (templaatti) on suorassa kosketuksessa vastaanottajamolekyylin (tuotteen) kanssa. Tässä mielessä RNA ei siis ole proteiinin templaatti, koska sanansaattaja-RNA:n ja proteiinin välillä ei tapahdu suoraa vuorovaikutusta translaation aikana (ks. Proteiinien biosynteesi). Tapauksissa, jotka ovat poikkeus keskeisestä dogmasta, viruksen RNA syntetisoidaan RNA:sta käyttäen RNA:ta templaattina (ks. RNA-virukset) ja lisäksi DNA syntetisoidaan käyttäen RNA:ta templaattina retrovirusten infektion aikana (ks. myös käänteinen transkriptio).
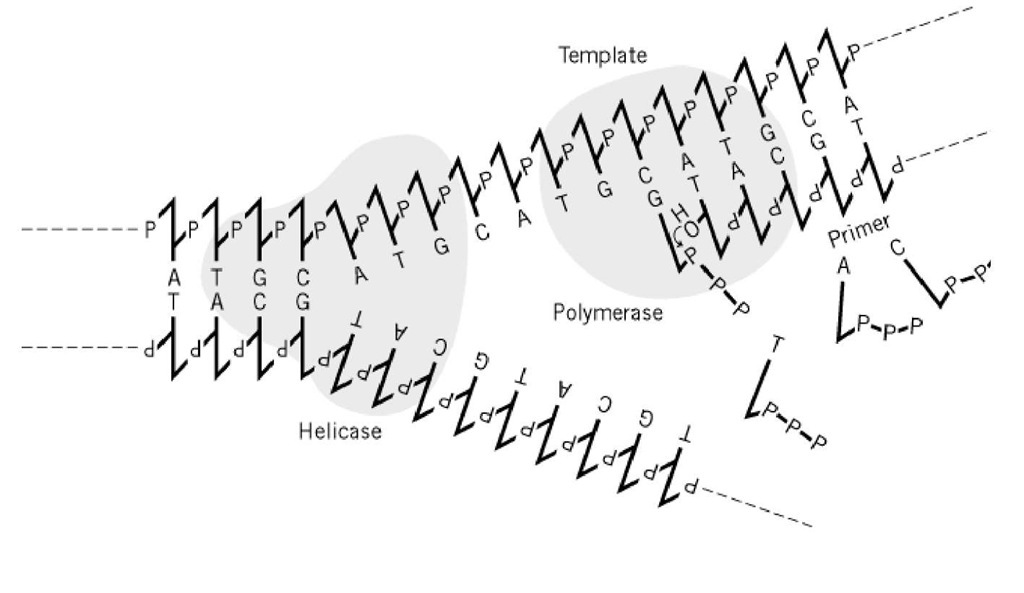
DNA:n replikaatiossa kaksoiskierre purkautuu, ja jokaista yksijuosteista DNA-molekyyliä käytetään mallina komplementaarisen juosteen syntetisoimiseksi. Koska DNA käy läpi puolikonservatiivisen replikaation, vanhempien DNA toimii mallina ja siitä tulee tytär-DNA-molekyylin osa. Transkriptiossa sitä vastoin DNA-juoste toimii mallina RNA:n synteesille, joka sitten dissosioituu mallista (ks. Transkriptio). DNA-polymeraasit, jotka kokoavat deoksiribonukleotideja templaatti-DNA-juosteeseen adeniini (A):tymiini (T), guaniini (G):sytosiini (C) -emäskomplementtisäännön mukaisesti (Watson-Crick-emäspariutuminen), tarvitsevat sekä templaatin että alukkeen, jotta reaktio voisi tapahtua (kuva 1). Näin ollen DNA-synteesiä edeltää pienten, alle 10 bp:n pituisten RNA-molekyylien synteesi RNA-primaasin avulla, joka voi käynnistää templaatti-DNA:n kanssa komplementaarisen RNA:n synteesin (ks. Okazaki Fragments). Alukkeena oleva RNA korvataan lopulta DNA:lla RNA-alukkeen synteesiä edeltävältä alueelta jatketun jäljessä olevan DNA-juosteen kautta (ks. Replikaatiohaarukka (Y-haarukan välivaihe)). A:n ja T:n sekä G:n ja C:n emäskomplementaarisuuden kautta syntyvä templaatti-tuotesuhde ei ole 100-prosenttisen tarkka, mutta se johtaa virheparin muodostumiseen kerran 10 3 – 104 kertaa. Toisaalta virhepareja syntyy yleensä vain kerran 10-10:stä, kun genomit replikoituvat solun sisällä. Tämä saavutetaan itse DNA-polymeraasien oikolukutoiminnalla ja muilla DNA:n korjausmekanismeilla.
Kuva 1. DNA-polymeraasi tarvitsee templaatin ja alukkeen. Kasvavassa replikaatiohaarukassa DNA-heliksaasin toimittamat yksittäiset DNA-säikeet toimivat templaaseina DNA-polymeraaseille. Entsyymit syntetisoivat fosfodiesterisidoksia alukkeen 3′( pään, vastasyntetisoidun säikeen ja deoksiribonukleotiditrifosfaatin välille, joka on emäspariutunut oikein templaattisäikeen kanssa. Tässä esitetään kaavamaisesti vain johtavan säikeen synteesi.
Templaatti-tuote-suhde on erittäin tärkeä geneettisen informaation siirrossa biologisissa järjestelmissä. Suhteen fysikaalis-kemiallinen luonne itsessään ei kuitenkaan ole riittävän tarkka tarkkaan säädeltyjen biologisten prosessien suorittamiseen, ja monet proteiinit on kehitetty korjaamaan väistämättä syntyviä virheitä.