
Szablon jest zdefiniowany w 1978 Webster’s New Collegiate Dictionary jako cząsteczka (taka jak RNA) w systemie biologicznym, która przenosi kod genetyczny dla innej cząsteczki. Również, 1995 Concise Oxford Dictionary, Ninth Edition opisuje szablon jako wzór molekularny rządzący montażem białka, itp. Zgodnie z tymi szerokimi definicjami, DNA jest szablonem dla DNA (w replikacji DNA) i dla RNA (w transkrypcji ), a RNA jest szablonem dla białka (w translacji ). Innymi słowy, szablon jest jednostką molekularną niezbędną do przeniesienia informacji genetycznej z DNA do DNA, z DNA do RNA do białka (główny dogmat). Dokładniej, słowo „szablon” jest używane, gdy cząsteczka przenosząca informację (szablon) jest w bezpośrednim kontakcie z cząsteczką odbiorcy (produktem). Zatem w tym sensie RNA nie jest szablonem dla białka, ponieważ podczas translacji nie dochodzi do bezpośredniej interakcji między mesenger RNA a białkiem (patrz biosynteza białka). W przypadkach, które są wyjątkiem od centralnego dogmatu, wirusowe RNA jest syntetyzowane z RNA przy użyciu RNA jako szablonu (zob. Wirusy RNA), a ponadto DNA jest syntetyzowane przy użyciu RNA jako szablonu podczas infekcji retrowirusów (zob. też Odwrotna transkrypcja).
W replikacji DNA, podwójna helisa jest rozwijana, a każda jednoniciowa cząsteczka DNA jest używana jako szablon do syntezy komplementarnej nici. Ponieważ DNA ulega replikacji półkonserwatywnej, DNA rodzicielskie służy jako szablon i staje się składnikiem cząsteczki DNA córki. W transkrypcji natomiast, nić DNA służy jako szablon do syntezy RNA, który następnie odłącza się od szablonu (patrz Transkrypcja). Polimerazy DNA, które montują deoksyrybonukleotydy na matrycy DNA zgodnie z zasadą komplementarności zasad Adenina (A):Tymina (T), Guanina (G):Cytozyna (C) (parowanie zasad Watsona-Cricka), wymagają do zajścia reakcji zarówno matrycy, jak i startera (Rys. 1). W konsekwencji synteza DNA poprzedzona jest syntez± małych cz±steczek RNA o masie mniejszej niż 10 bp przez primazę RNA, która może zainicjować syntezę RNA komplementarnego do szablonowego DNA (patrz Fragmenty Okazaki). Pryma RNA jest ostatecznie zastępowana przez DNA za pośrednictwem nici DNA przedłużonej z regionu poprzedzającego syntezę primera RNA (patrz: Widełki replikacyjne (Y-Fork Intermediate)). Zależność szablon-produkt poprzez komplementarność zasad A do T i G do C nie jest w 100% dokładna, ale powoduje powstawanie błędnych par raz na 10 3 do 104 razy. Z drugiej strony, podczas replikacji genomów w komórce, błędne pary występują na ogół tylko raz na 10 do 10 . Osiąga się to dzięki aktywności korekcyjnej samych polimeraz DNA oraz innym mechanizmom naprawy DNA.
Rysunek 1. Polimeraza DNA wymaga szablonu i primera. Na rosnących widełkach replikacyjnych pojedyncze nici DNA dostarczane przez helikazy DNA służą jako szablony dla polimeraz DNA. Enzymy te syntetyzują wiązania fosfodiestrowe pomiędzy 3′( końcem primera, nowo syntetyzowanej nici, a deoksyrybonukleotydem-trójfosforanem, który jest prawidłowo sparowany zasadami z nicią szablonu. Schematycznie pokazana jest tu tylko synteza nici wiodącej.
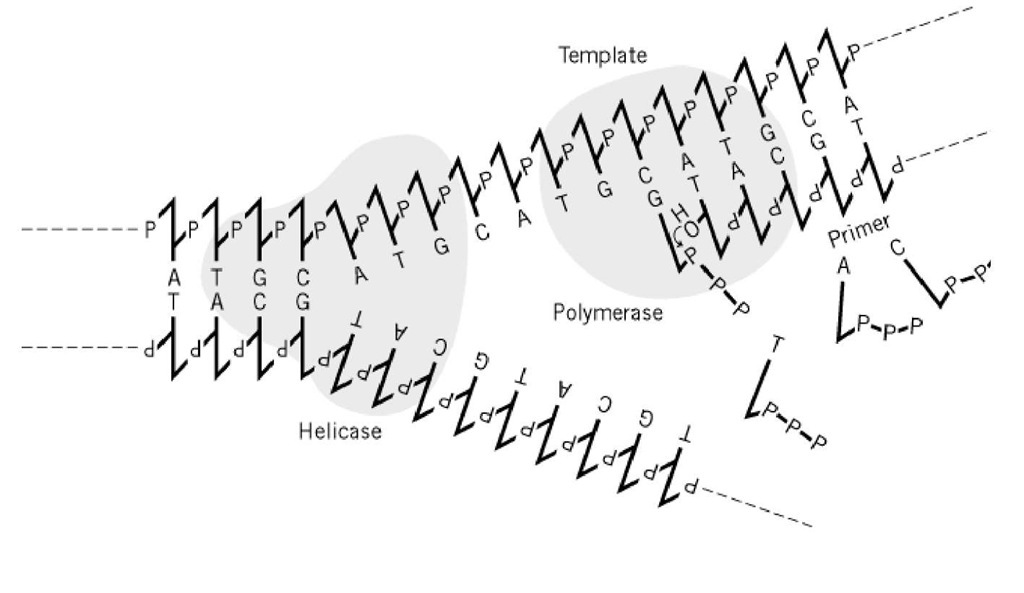
Zależność szablon-produkt jest bardzo ważna w przekazywaniu informacji genetycznej w systemach biologicznych. Jednak sama fizyko-chemiczna natura tej relacji nie jest wystarczająco dokładna, aby wykonywać precyzyjnie regulowane procesy biologiczne, a wiele białek zostało wyewoluowanych w celu naprawy błędów, które nieuchronnie powstają.
.