
Introducerea ciclinei
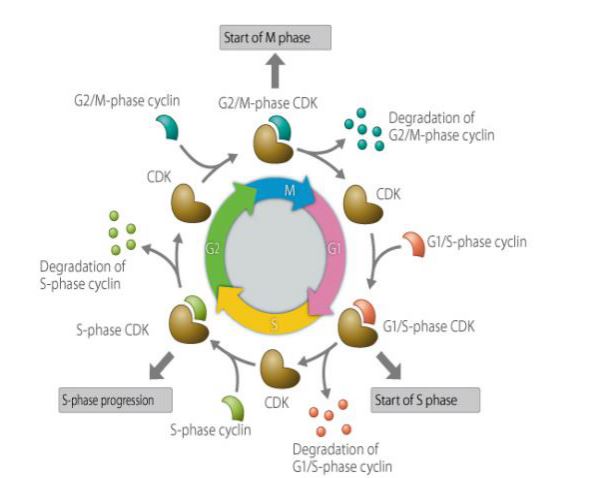
Ciclina este un tip de proteină care se exprimă, se acumulează și se descompune în ciclul celular și interacționează cu kinazele dependente de ciclină pentru a afecta funcția ciclului celular. Ciclina este implicată în proteinele reglate de ciclul celular, iar concentrația sa este ciclică și ciclică în ciclul celular. În funcție de stadiul ciclului celular, uneori concentrația este de câteva mii de ori mai mare, iar alteori scade la zero. În calitate de subunitate de reglare, ciclina se leagă și activează protein-kinazele dependente de ciclină. Diferite cicline sunt exprimate în diferite perioade din ciclul celular. În celulele mamiferelor, ciclina A începe să se exprime și să se acumuleze treptat în faza G1 timpurie, ajungând la joncțiunea G1/S, iar conținutul atinge maximul și rămâne în faza G2/M. Ciclina B se exprimă de la sfârșitul fazei G1 și se acumulează treptat. Ea atinge maximul în faza G2 târzie și se menține până la jumătatea fazei M, iar apoi se degradează rapid. Ciclina D, care este o ciclină din faza G1, este exprimată continuu în ciclul celular, în timp ce ciclina E începe să se exprime și să se acumuleze treptat în faza M târzie și în faza G1 timpurie, atingând maximul în faza G1 târzie, iar apoi își atinge treptat conținutul maxim, scăzând până în faza G2 târzie. Ciclina joacă un rol important în reglarea ciclului celular. Studierea mecanismului ciclinei are o mare importanță pentru tratamentul cancerului.
Membrii familiei ciclinei și, respectiv, funcțiile lor
Până în prezent, ciclinele umane au fost izolate și denumite într-un total de 8 categorii, și anume de la A la H, și includ unele subclase (cum ar fi ciclinele D1, D2, D3), dintre care F, G, H sunt cele mai recente descoperiri. Toate ciclinele au anumite similitudini de secvență de aminoacizi și, astfel, servesc drept markeri structurali moleculari ai ciclinei. Această regiune de aminoacizi omologi din cicline este așa-numita casetă a ciclinei, care este o regiune de secvență de 100-150 de aminoacizi foarte conservată între ciclinele diferitelor organisme. Ciclina umană conține, de asemenea, o casetă defectă sau o regiune de secvență de aminoacizi bogată în reziduuri de prolină, acid glutamic, acid aspartic, serină și treonină (denumită în continuare regiunea de secvență de aminoacizi PEST). Legarea ciclinei B de kinaza dependentă de cicline p34CDC2 controlează intrarea și ieșirea din mitoză, care se sintetizează din faza S în faza G2 /M și se acumulează treptat și se leagă de p34CDC2. În faza M, aceasta se degradează treptat; dimpotrivă, ieșirea din faza M depinde de degradarea rapidă a ciclinei în faza târzie a diviziunii, ceea ce duce la eliberarea subunității inactive p34CDC2. În celulele umane, ciclina B se acumulează în citoplasmă și intră în nucleu înainte de dispariția membranei nucleare. Odată activată, ciclina B/CCDC fosforilează un grup de proteine, inclusiv lamin, vimentină și calciu, care joacă un rol important în concentrarea cromozomilor, dezintegrarea membranei nucleare, depolimerizarea filamentelor intermediare și reorganizarea microfilamentelor. Ciclina B joacă, de asemenea, un rol important în tranziția de fază a mitozei, care este o proteină clasică a ciclului de clivaj. Ciclina A joacă un rol atât în tranzițiile de fază G2/M, cât și în G1/S. Apare înainte de începerea sintezei ADN-ului și crește treptat până la etapa de pre-scindare și se degradează în etapa de mijloc. Atunci când induce mitoza, aceasta diferă de adevărata ciclină B mitotică în multe aspecte. Conținutul de ciclină C se modifică puțin de-a lungul ciclului celular, cu doar o ușoară creștere la începutul G1. ARNm al ciclinei C atinge vârful la mijlocul lui G1 în celulele HeLa sincronizate, ceea ce este mai devreme decât ciclina A, care poate avea un rol în G1. Cyclin E are o expresie periodică distinctă în ciclul celular, iar vârful său este în tranziția de fază G1 /S. Aceasta are rolul de a controla intrarea celulei în faza S, care poate fi factorul care limitează rata de tranziție în faza G1 /S. În celulele mamiferelor, la fel ca și ciclina A, se leagă de p33CDK2, un complex care are activitate de kinază a histonei H1 în faza timpurie G1 și S, dar ciclina A și E pot interacționa cu proteina retinoblastomului pentru a separa complexele, ceea ce sugerează că complexele CDK2/ciclină A sau E pot regla indirect expresia genică în fazele G1 și S. Ciclina E poate juca un rol în ciclina D, care joacă un rol important în inițierea replicării ADN, în conversia G1 /S, iar supraexprimarea acesteia accelerează intrarea celulei în faza S. Ciclina D poate controla faza G1 timpurie a procesului, fie înainte, fie în același timp cu ciclina E. Ciclina D are trei subtipuri (D1, D2, D3), iar expresia sa variază de la o celulă la alta și este controlată de diferite regiuni cromozomiale (D1: 11q13; D2: 12p13; D3: 6p21). Subtipurile pot avea efecte diferite. Caracteristicile remarcabile ale ciclinei D sunt: este indusă să se exprime ca răspuns la stimuli externi, prezentând o inductibilitate ridicată a factorilor de creștere, acționând ca un senzor al factorilor de creștere, care joacă un rol important în corelarea semnalelor externe cu ciclul celular intern. Rolul, la rândul său, a dedus că exprimarea sa necontrolată face ca ciclul celular să nu mai depindă sau să fie mai puțin dependent de factorii de creștere și poate induce cancerul. Complexul ciclină D-CDK este cel mai bun candidat pentru proteina-kinaza retinoblastomului (Rb) în stadiul G1. Ciclina D se leagă de regiunea N-terminală a proteinei Rb, fosforilează Rb în faza G1 târzie, iar sinteza și activarea ciclinei D1 duce la inactivarea fosforilării Rb și la reglarea în jos a ciclinei D1, iar complexul D1-CDK4 formează un feedback negativ în faza G1, care, la rândul său, oprește expresia ciclinei D1. Ciclina D și Rb joacă un rol important în proliferarea și diferențierea celulară, atunci când ciclina D1 este diferită în faza G1. În expresia sursei, fosforilarea Rb este mai timpurie decât în mod normal; faza G1 este accelerată, iar anticorpul anticiclin D1 este microinjectat în G1 devreme în celulele aflate în metafază; majoritatea celulelor sunt arestate înainte de faza S, iar lipsa de Rb funcțional nu poate cauza acest blocaj; se concluzionează că rolul important al ciclinei D1 este de a inactiva fosforilarea lui Rb, inducând astfel intrarea în faza S și replicarea ADN-ului. CyclinF este cea mai abundentă proteină ciclină (greutate moleculară 87 kDa), iar ARNm al acesteia este universal exprimat în diferite celule de țesut uman și există schimbări semnificative în ciclul celular. Vârful este în faza G2, la fel ca și în cazul ciclinei A, și scade înainte ca nivelul ARNm al ciclinei B să scadă. Proteina ciclină F se acumulează în faza intercelulară și este distrusă în timpul fazei de mitoză. Aceasta se află în nucleu în majoritatea celulelor. Supraexprimarea sau mutația în celulele umane duce la o lipsă a regiunilor secvenței de aminoacizi PEST, ceea ce duce la o creștere a celulelor în faza G2. Ciclina G seamănă cel mai mult cu ciclina B din drojdia de fisiune, care joacă un rol în conversia G1/S, dar ARNm al ciclinei G nu are o dependență celulară evidentă, dar poate fi stimulată de creșterea celulară. Factorul induce și menține un nivel ridicat. Gena cyclin g conține două situsuri diferite de legare a p53, dintre care unul se află în amonte de situsul de inițiere a transcripției, ceea ce sugerează că p53 are potențialul de a activa eficient gena cyclin g. Ciclina H, care este un mediator în aval al p53, cel puțin în ceea ce privește efectul biologic, este o proteină cu o greutate moleculară de 37 kD descoperită prin izolarea și purificarea kinazei activatoare a CDK (CAK).
Funcția ciclinei
Studiul relației dintre ciclul celular și cancer are o semnificație orientativă pentru cercetarea clinică oncologică. Poate oferi anumite ținte pentru tratamentul clinic, cum ar fi blocarea ciclinei D sau imitarea acțiunii inhibitorilor ciclului celular pentru a inhiba diviziunea celulelor canceroase, și poate oferi anumiți indicatori pentru diagnosticul clinic, diagnosticul diferențial și prognosticul. Michaelet al a analizat rearanjarea genei bcl-1 și expresia proteinei ciclinei D1 în 32 de cazuri de limfom cu celule de manta, 17 cazuri de rearanjare a genei bcl-1 și 24 de cazuri de expresie citoplasmatică a ciclinei D1, iar 40 de cazuri din grupul de control doar 9 cazuri de limfom cu celule B non-MCL au fost pozitive. Aceștia consideră că ciclina D1 poate fi utilizată ca diagnostic diferențial al MCL și diagnostic diferențial al limfomului cu celule B. Ciclina este exprimată periodic în ciclul celular, iar anumiți complecși specifici ciclină-CDK sunt necesari pentru trecerea printr-un anumit ciclu celular, sugerând că ciclina poate fi utilizată ca indicator al stării proliferative a celulelor. Raportul de expresie al unei anumite cicline poate prezice proporția de celule dintr-un anumit țesut care se află într-un anumit ciclu celular. Cu cât gradul de malignitate al celulelor tumorale este mai ridicat, cu atât mai gravă este tulburarea ciclului celular, astfel încât este posibil să se reflecte prognosticul pacienților prin intermediul informațiilor prezentate în ciclul celular și să acționeze ca un posibil indicator de prognostic. Keyomarsi et al. au folosit 9 cazuri de probe chirurgicale de cancer de sân și țesuturi necanceroase adiacente ca studii de control: 8 cazuri de țesuturi canceroase cyclin E au fost exprimate anormal din punct de vedere calitativ și cantitativ și au existat cel puțin 3 greutăți moleculare diferite. Ciclina E a fost supraexprimată, în timp ce c-erbB2 este supraexprimată doar în 3 cazuri. Odată cu creșterea stadializării tumorii, conținutul de proteină ciclină E a crescut semnificativ, în timp ce nivelul antigenului nuclear al celulelor proliferante (PCNA) a crescut doar puțin. Structura cyclin E a fost observată în 4 dintre cele mai înalte grade de stadializare. Dutta et al. au folosit anticorpi anticiclin A, B, E pentru a detecta 48 de secțiuni de parafină de cancer de sân și au constatat că indicele mediu pozitiv al ciclinei A și B a crescut semnificativ odată cu creșterea fazei S (P < 0,05). Ciclina A a fost corelată pozitiv cu Ki-67 și cu fracția de fază S (P < 0,05). Bellacosaet al au studiat 51 de cazuri de carcinom primar cu celule scuamoase laringian și au urmărit timp de 29 de luni. Se consideră că amplificarea genei ciclinei poate fi utilizată ca indicator independent de prognostic pentru cancerul laringian. Cu toate acestea, Bettiche et al. au studiat 53 de cazuri de cancer pulmonar cu celule non-mici cu rezecție chirurgicală, iar 25 de cazuri prezentau supraexpresia ciclinei D1, care a fost asociată cu o diferențiere tisulară slabă, mai puțină infiltrație limfocitară în tumoră și o rată de recidivă locală mai mică (P < 0. 05). Aceștia consideră că sunt necesare studii clinice mai ample pentru a testa în continuare semnificația prognostică a expresiei ciclinei D1.
Referință
- He G, Kuang J, Koomen J, et al. Recrutarea antigenului nuclear trimeric al celulelor proliferante de către kinazele dependente de cicline în faza G1 în urma deteriorării ADN-ului cu agenți antitumorali pe bază de platină. British Journal of Cancer. 2013, 109(9):2378-2388.
- Gang W, Cao J, Chen P, et al. Temporal and Spatial Expression of Cyclin H in Rat Spinal Cord Injury. Neuromolecular Medicine. 2011, 13(3):187-196.
- Ibrahim N, Nazimi A J, Ajura A J, et al. The Clinical Features and Expression of bcl-2, Cyclin D1, p53, and Proliferating Cell Nuclear Antigen in Syndromic and Nonsyndromic Keratocystic Odontogenic Tumor. Journal of Craniofacial Surgery. 2016, 27(5):1.
- Wang X, Wolgemuth D J, Baxi L V. Supraexpresia HOXB5, a ciclinei D1 și a PCNA în malformația adenomatoidă chistică congenitală. Diagnostic fetal & Terapie. 2011, 29(4):315-320.