
Un modèle est défini dans le Webster’s New Collegiate Dictionary de 1978 comme une molécule (comme l’ARN) dans un système biologique qui porte le code génétique d’une autre molécule. De même, le Concise Oxford Dictionary de 1995, neuvième édition, décrit un modèle comme le schéma moléculaire régissant l’assemblage d’une protéine, etc. Selon ces définitions générales, l’ADN est le modèle de l’ADN (dans la réplication de l’ADN) et de l’ARN (dans la transcription), et l’ARN est le modèle de la protéine (dans la traduction). En d’autres termes, une matrice est une entité moléculaire essentielle au transfert de l’information génétique de l’ADN à l’ARN et à la protéine (le dogme central). Plus précisément, le mot « template » est utilisé lorsqu’une molécule transférant l’information (le template) est en contact direct avec la molécule réceptrice (le produit). Par conséquent, en ce sens, l’ARN n’est pas un modèle pour les protéines car aucune interaction directe entre l’ARN messager et la protéine ne se produit au cours de la traduction (voir Biosynthèse des protéines). Dans les cas qui font exception au dogme central, l’ARN viral est synthétisé à partir de l’ARN en utilisant l’ARN comme matrice (voir Virus à ARN) et, de plus, l’ADN est synthétisé en utilisant l’ARN comme matrice lors de l’infection des rétrovirus (voir aussi Transcription inverse).
Dans la réplication de l’ADN, la double hélice est déroulée, et chaque molécule d’ADN simple brin est utilisée comme matrice pour synthétiser un brin complémentaire. Comme l’ADN subit une réplication semi-conservative, l’ADN parental sert de matrice et devient un composant de la molécule d’ADN fille. Dans la transcription, en revanche, un brin d’ADN sert de matrice pour la synthèse de l’ARN qui se dissocie ensuite de la matrice (voir Transcription). Les ADN polymérases qui assemblent les désoxyribonucléotides sur le brin d’ADN matrice selon la règle de complémentarité des bases Adénine (A):Thymine (T), Guanine (G):Cytosine (C) (appariement des bases de Watson-Crick) ont besoin à la fois d’une matrice et d’une amorce pour que la réaction ait lieu (Fig. 1). Par conséquent, la synthèse de l’ADN est précédée par la synthèse de petites molécules d’ARN de moins de 10 pb par l’ARN primase, qui peut initier la synthèse de l’ARN complémentaire de l’ADN matrice (voir Fragments d’Okazaki). L’ARN amorce est finalement remplacé par de l’ADN grâce au brin d’ADN retardé qui s’étend à partir de la région précédant la synthèse de l’ARN amorce (voir Fourche de réplication (intermédiaire Y-Fork)). La relation modèle-produit par le biais de la complémentarité des bases de A à T et de G à C n’est pas exacte à 100 %, mais aboutit à la formation d’un mauvais appariement une fois tous les 10 3 à 104 fois. En revanche, les erreurs d’appariement ne se produisent généralement qu’une fois sur 10 à 10 lorsque les génomes sont répliqués dans la cellule. Ceci est obtenu par l’activité de relecture des ADN polymérases elles-mêmes et par d’autres mécanismes de réparation de l’ADN.
Figure 1. L’ADN polymérase nécessite une matrice et une amorce. Au niveau d’une fourche de réplication en croissance, des brins simples d’ADN fournis par l’ADN hélicase servent de tempes aux ADN polymérases. Les enzymes synthétisent des liaisons phosphodiester entre l’extrémité 3′( de l’amorce, brin nouvellement synthétisé, et le désoxyribonucléotide-triphosphate qui est correctement apparié en base avec le brin matrice. Seule la synthèse du brin d’attaque est représentée ici de façon schématique.
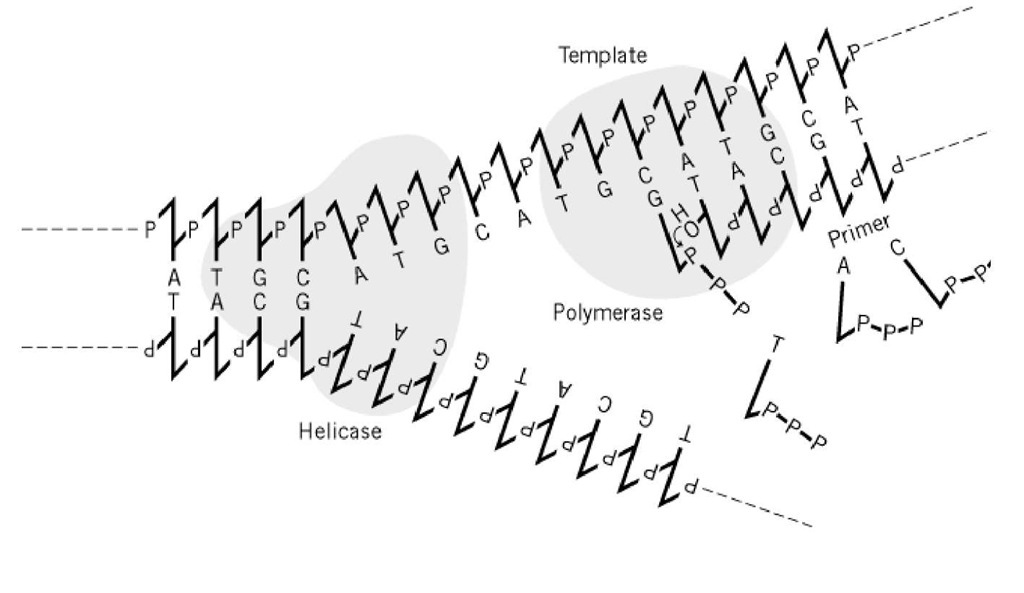
La relation matrice-produit est très importante dans le transfert de l’information génétique dans les systèmes biologiques. Cependant, la nature physico-chimique de la relation elle-même n’est pas suffisamment précise pour réaliser les processus biologiques régulés avec précision, et de nombreuses protéines ont été développées pour réparer les erreurs qui en résultent inévitablement.