
Indledning om cyklin
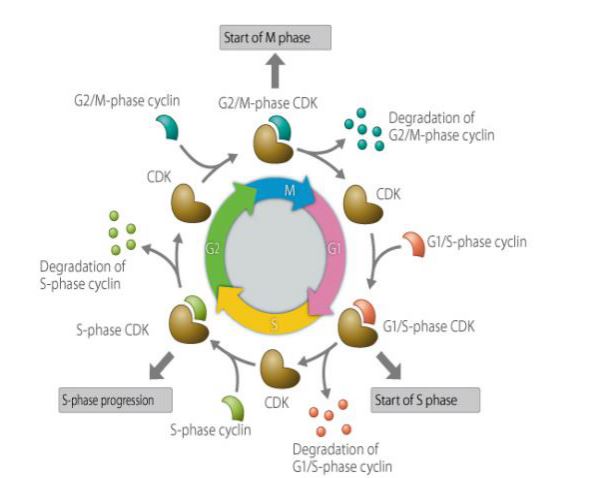
Cyclin er en type protein, der udtrykkes, akkumuleres og nedbrydes i en cellecyklus, og det interagerer med cyklinafhængige kinaser for at påvirke cellecyklusfunktionen. Cyclin har involveret i cellecyklusregulerede proteiner, og dets koncentration er cyklisk og cyklisk i cellecyklusen. Afhængigt af stadiet i cellecyklussen er koncentrationen nogle gange så høj som flere tusinde gange, og nogle gange falder den til nul. Som en regulerende underenhed binder cyklin til og aktiverer cyclinafhængige proteinkinaser. Forskellige cykliner udtrykkes i forskellige perioder i cellecyklusen. I pattedyrceller begynder cyklin A at udtrykke sig og akkumuleres gradvist i den tidlige G1-fase og når G1/S-overgangen, og indholdet når sit maksimum og forbliver i G2/M-fasen. Cyclin B udtrykkes fra den sene G1-fase og akkumuleres gradvist. Det når sit maksimum i den sene G2-fase og opretholdes til midten af M-fasen, hvorefter det hurtigt nedbrydes. Cyklin D, som er et cyklin fra G1-fasen, udtrykkes kontinuerligt i cellecyklusen, mens cyklin E begynder at udtrykke sig og gradvist ophobes i den sene M-fase og den tidlige G1-fase, når maksimum i den sene G1-fase, og når derefter gradvist sit maksimale indhold og aftager indtil den sene G2-fase. Cyclin spiller en vigtig rolle i reguleringen af cellecyklussen. Studiet af cyklins mekanisme har stor betydning for behandlingen af kræft.
Cyclin-familiemedlemmer og deres funktioner henholdsvis
Der er indtil nu blevet isoleret og navngivet menneskelige cykliner i i alt 8 kategorier, nemlig A til H, og omfatter nogle underklasser (såsom cyclin D1, D2, D3), hvoraf F, G, H er de seneste opdagere. Alle cyklinerne har visse ligheder i aminosyresekvensen og tjener således som molekylære strukturelle markører for cykliner. Denne homologe aminosyreregion i cykliner er den såkaldte cyklinkassette, som er en meget bevaret 100-150 aminosyre-sekvensregion mellem cykliner fra forskellige organismer. Det menneskelige cyklin indeholder også en defekt kassette eller en aminosyresekvensregion, der er rig på prolin-, glutaminsyre-, asparaginsyre-, serin- og threoninrester (i det følgende benævnt “PEST-aminosyresekvensregionen”). Cyklin B’s binding af cyklin B til den cyklinafhængige kinase p34CDC2 styrer indgangen til og udgangen af mitose, som syntetiseres fra S-fasen til G2 /M-fasen og gradvist akkumuleres og binder sig til p34CDC2. I M-fasen nedbrydes den gradvist; tværtimod afhænger det at forlade M-fasen af den hurtige nedbrydning af cyklinen i den sene fase af divisionen, hvilket resulterer i frigivelse af den inaktive underenhed p34CDC2. I humane celler ophobes cyklin B i cytoplasmaet og kommer ind i kernen, før kerneembranen forsvinder. Når cyklin B/CDC er aktiveret, fosforylerer cyklin B/CDC en gruppe proteiner, herunder lamin, vimentin og calcium, som spiller en vigtig rolle i kromosomkoncentration, opløsning af kernemembranen, depolymerisering af intermediære filamenter og reorganisering af mikrofilamenter. Cyclin B spiller også en vigtig rolle i faseovergangen i mitose, som er et klassisk spaltningscyklusprotein. Cyclin A spiller en rolle i både G2/M- og G1/S-faseovergangene. Det optræder før starten af DNA-syntesen, og det stiger gradvist indtil præ-spaltningsstadiet og nedbrydes i mellemstadiet. Når det inducerer mitose, adskiller det sig fra det ægte mitotiske cyklin B i mange henseender. Cyclin C-indholdet ændrer sig kun lidt gennem hele cellecyklussen, med kun en lille stigning i det tidlige G1. Cyclin C mRNA topper midt i G1 i synkroniserede HeLa-celler, hvilket er tidligere end cyclin A, som kan have en rolle i G1. Cyclin E har en tydelig periodisk ekspression i cellecyklusen, og dets højdepunkt ligger i overgangen mellem G1 og S-fasen. Det har den funktion at kontrollere, at cellen går ind i S-fasen, hvilket kan være den hastighedsbegrænsende faktor i G1/S-faseovergangen. I pattedyrceller binder det ligesom cyklin A til p33CDK2, et kompleks, der har histon H1-kinaseaktivitet i tidlig G1- og S-fase, men cyklin A og E kan interagere med retinoblastoma-protein for at adskille komplekser, hvilket tyder på, at CDK2/cyklin A eller E-komplekser indirekte kan regulere genekspression i G1- og S-faser. Cyclin E kan spille en rolle i cyclin D, som spiller en vigtig rolle i initiering af DNA-replikation, G1 /S-konvertering, og dets overekspression fremskynder cellens indtræden i S-fasen. Cyclin D kan kontrollere den tidlige G1-fase af processen, enten før eller samtidig med cyclin E. Cyclin D har tre undertyper (D1, D2, D3), og dets udtryk varierer fra celle til celle og kontrolleres af forskellige kromosomale regioner (D1: 11q13; D2: 12p13; D3: 6p21). Undertyperne kan have forskellige virkninger. Cyklin D’s bemærkelsesværdige kendetegn er, at det induceres til at udtrykke sig som reaktion på eksterne stimuli, idet det udviser en høj inducerbarhed for vækstfaktorer og fungerer som en vækstfaktorsensor, der spiller en vigtig rolle ved at forbinde eksterne signaler med den indre cellecyklus. Den rolle, til gengæld udledes dens ukontrollerede udtryk gør cellecyklusen ikke længere eller mindre afhængig af vækstfaktorer og kan fremkalde kræft. Cyklin D-CDK-komplekset er den bedste kandidat til G1-stadiet retinoblastoma (Rb)-proteinkinase. Cyklin D binder sig til den N-terminale region af Rb-protein, fosforylerer Rb i den sene G1-fase, og cyklin D1-syntese og -aktivering fører til inaktivering af Rb-fosforylering og nedregulering af cyklin D1, og D1-CDK4-komplekset danner en negativ feedback i G1-fasen, som igen lukker ned for ekspressionen af cyklin D1. Cyclin D og Rb spiller en vigtig rolle i celleproliferation og differentiering, når cyclin D1 er anderledes i G1-fasen. I kildeudtrykket er Rb-fosforylering tidligere end normalt; G1-fasen accelereres, og anti-cyclin D1-antistof mikroinjiceres i G1 tidligt til metafaseceller; de fleste celler arresteres før S-fasen, og mangel på funktionel Rb kan ikke forårsage denne blokering; det konkluderes, at cyclin D1’s vigtige rolle er at inaktivere fosforyleringen af Rb og derved inducere indgangen til S-fasen og DNA-replikation. CyclinF er det hyppigst forekommende cyklinprotein (molekylvægt 87 kDa), og dets mRNA er universelt udtrykt i forskellige humane vævsceller, og der er betydelige ændringer i cellecyklusen. Toppen er i G2-fasen, ligesom cyklin A, og falder, før cyklin B mRNA-niveauet falder. Cyclin F-proteinet ophobes i den intercellulære fase og ødelægges i mitosefasen. Det befinder sig i kernen i de fleste celler. Overekspression eller mutation i menneskelige celler resulterer i en mangel på PEST-aminosyresekvensregioner, hvilket fører til en stigning i G2-faseceller. Cyclin G ligner mest Cyclin B fra fissionsgær, som spiller en rolle i G1/S-konverteringen, men mRNA af cyclin G har ingen tydelig celleafhængighed, men kan stimuleres af cellevækst. Faktoren inducerer og opretholder et forhøjet niveau. Cyclin g-genet indeholder to forskellige p53-bindingssteder, hvoraf det ene ligger opstrøms for transkriptionsinitieringsstedet, hvilket tyder på, at p53 har potentiale til effektivt at aktivere cyclin g-genet. Cyclin H, som er en downstream mediator for p53 i det mindste i biologisk effekt, er et protein med en molekylvægt på 37 kD, der er fundet ved isolering og oprensning af CDK-aktiverende kinase (CAK).
Funktion af cyclin
Undersøgelsen af forholdet mellem cellecyklus og kræft har vejledende betydning for den kliniske onkologiske forskning. Det kan give visse mål for klinisk behandling, f.eks. blokering af cyklin D, eller efterligne virkningen af cellecyklusinhibitorer for at hæmme kræftcelledelingen og give nogle indikatorer for klinisk diagnose, differentialdiagnose og prognose. Michaelet al analyserede bcl-1 genomlægning og cyclin D1 proteinudtryk i 32 tilfælde af mantelcellelymfom, 17 tilfælde af bcl-1 genomlægning og 24 tilfælde af cytoplasmatisk cyclin D1-ekspression, og 40 tilfælde af kontrolgruppe kun 9 tilfælde af ikke-MCL B-celle lymfom var positive. De mener, at cyklin D1 kan anvendes som differentialdiagnose af MCL og som differentialdiagnose af B-celle lymfom. Cyclin udtrykkes periodisk i cellecyklusen, og visse specifikke cyclin-CDK-komplekser er nødvendige for passage gennem en bestemt cellecyklus, hvilket tyder på, at cyclin kan bruges som en indikator for cellers proliferative tilstand. Ekspressionsforholdet for et bestemt cyklin kan forudsige andelen af celler i et bestemt væv, der befinder sig i en bestemt cellecyklus, i et bestemt væv. Jo mere maligne tumorcellerne er, jo mere alvorlig er cellecyklusforstyrrelsen, så det er muligt at afspejle patienternes prognose gennem de oplysninger, der vises i cellecyklusen, og fungere som en mulig prognostisk indikator. Keyomarsi et al. anvendte 9 tilfælde af kirurgiske prøver af brystkræft og tilstødende ikke-cancerøse væv som kontrolundersøgelser: I 8 tilfælde af kræftvæv var cyklin E unormalt udtrykt i kvalitet og kvantitet, og der fandtes mindst 3 forskellige molekylære vægte. Cyclin E var overeksprimeret, mens c-erbB2 kun var overeksprimeret i 3 tilfælde. Med stigningen i tumorstadiet steg indholdet af cyklin E-protein betydeligt, mens niveauet af PCNA (proliferating cell nuclear antigen) kun steg svagt. Cyclin E-strukturen blev observeret i 4 af de højeste stadieinddelingsklasser. Dutta et al. anvendte anti-cyclin A, B, E antistoffer til påvisning af 48 paraffinsnit af brystkræft og fandt, at det gennemsnitlige positive indeks for cyclin A og B steg signifikant med stigningen i S-fasen (P < 0,05). Cyclin A var positivt korreleret med Ki-67 og med S-fasefraktionen (P < 0,05). Bellacosaet al undersøgte 51 tilfælde af primær larynxpladecellekarcinom i larynx og fulgte op i 29 måneder. Man mener, at cyklin-genamplifikation kan anvendes som en uafhængig prognostisk indikator for larynxkræft. Bettiche et al. undersøgte imidlertid 53 tilfælde af ikke-småcellet lungekræft med kirurgisk resektion, og 25 tilfælde havde cyclin D1-overekspression, hvilket var forbundet med dårlig vævsdifferentiering, mindre lymfocytinfiltration i tumoren og lavere lokal recidivrate (P < 0. 05). De mener, at der er behov for større kliniske undersøgelser for yderligere at teste den prognostiske betydning af cyclin D1-ekspression.
Reference
- He G, Kuang J, Koomen J, et al. Rekruttering af trimerisk proliferating cell nuclear antigen af G1-fase cyclinafhængige kinaser efter DNA-skader med platinbaserede antitumormidler. British Journal of Cancer. 2013, 109(9):2378-2388.
- Gang W, Cao J, Chen P, et al. Temporal and Spatial Expression of Cyclin H in Rat Spinal Cord Injury (Tidsmæssig og rumlig ekspression af Cyclin H i rygmarvsskader hos rotter). Neuromolekylær medicin. 2011, 13(3):187-196.
- Ibrahim N, Nazimi A J, Ajura A J, et al. De kliniske træk og ekspression af bcl-2, Cyclin D1, p53 og Proliferating Cell Nuclear Antigen i syndromisk og ikke-syndromisk keratocystisk odontogen tumor. Journal of Craniofacial Surgery. 2016, 27(5):1.
- Wang X, Wolgemuth D J, Baxi L V. Overekspression af HOXB5, cyclin D1 og PCNA i medfødt cystisk adenomatoid misdannelse. Fosterdiagnostik & Terapi. 2011, 29(4):315-320.